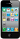
Таблица 1. Cостав спиртовой части нейтральных алкил-глицеридов из природных источников.
|
Спиртовый компонент |
Костный мозг человека |
Молоко человека |
Жир Гренландской акулы |
Кальмар |
|
14:0 |
|
|
2.0 |
0,7 |
|
15:0а |
|
|
0,7 |
0,3 |
|
16:0 |
29,4 |
23,9 |
9,1 |
58,3 |
|
16:1 |
|
Следы |
10:8 |
0,5 |
|
17:0а |
7,6 |
3,6 |
3,6 |
1,9 |
|
18:0 |
24,6 |
22,8 |
2,8 |
7,6 |
|
18:1 |
16,7 |
33,8 |
59,4 |
23,2 |
|
18:2 |
|
1,4 |
1,6 |
|
|
19:0а |
6,1 |
2,4 |
1,5 |
1,1 |
|
20:0 |
2,9 |
1,6 |
|
|
|
20:1 |
3,2 |
2,3 |
6,2 |
3,4 |
|
22:0 |
0,7 |
0,7 |
|
|
|
22:1 |
5,1 |
3,4 |
2,2 |
|
|
24:0 |
|
2,1 |
|
|
Биосинтез и распределение в животном мире
Биосинтез простых эфиров глицерина был исследован, главным образом на клетках млекопитающих и приведен на Рисунке 2. Исходным соединением для биосинтеза de novo является дигидроацетон фосфат (ДГАФ), который ацилируется acyl-CoA с ферментом ДГАФ ацилтрансферазой, в пероксисомах в sn-1 положении глицерина. Затем происходит замена ацильного радикала на алкильную группу пероксисомальным мембрансвязанным ферментом (de Vet et al., 2000). Далее, кето группа ДГАФ восстанавливается до гидроксильной с образованием гидроксильной группы в sn-2 положении глицерина с последующим ацилированием с acyl-CoA. Дальнейшая трансформация 1-O-алкил-2-ацил-глицерофосфата осуществляемая ферментом фосфогидролазой, при этом образуется ключевой продукт в цепи биосинтеза как нейтральных, так и полярных алкильных производных (Paltauf, 1983). 1-0-алкил-2-ацил-глицерин может метаболизировать по 2 механизмам: образовывать нейтральные АДАГ и под действием фермента этаноламин-фосфотрансферазы с кофактором цитидилдифосфо-этаноламином образуются фосфор-содержащие плазманил-этаноламины (Snyder, 1999). Последний, под действием Δ1-десатуразы с цитохромом b5 превращается в плазменил-этаноламины или плазмалогены (Brites et al., 2004). Тем не менее, биосинтез плазмалеген-холина до настоящего времени не ясен, хотя радиоизотопные исследования показали, что плазмалеген-холин является результатом превращения плазмалеген-этаноламина. Как полагают, это происходит под действием мембрансвязанной фосфолипазы D, которая может проявлять и трансферазную активность, обменивая этанолоамин в молекуле этаноламин-плазмалогене на холин.
Состав спиртовой группы в АДАГ был подробно исследован в жире печени акул семейства Squaliformes, в липидах телеостов, морских беспозвоночных и в клетках злокачественных опухолевых тканей. В Таблице 1 показан состав спиртовой группы в основных природных источниках. Спиртовые радикалы, как правило, насыщенные или мононенасыщены с числом атомов углерода от С-14 до С-22 составляют более 90% от суммы спиртов в АДАГ. Нечетные, полиненасыщенные и разветвленные спирты встречаются в незначительных количествах. Алкилированные радикалы короче С-14 найдены только у некоторых рыб в количествах менее 2%, в то время как С-24 спирты были найдены в молоке и костном мозге человека. Диеновые АДАГ, имеют главным образом морское происхождение. Разветвленные алкильные цепи, распространены в молоке жвачных животных (Hallgren et al., 1974b), где они составляют приблизительно 10% от суммы всех спиртов. Нечетные спирты (до 32%), включая 21% 1-O-pentadecyl глицерин были найдены у планктонного организма Clione limacina (Phleger et al., 1997). Самым распространенным в жире печени глубоководных акул (Deprez et al. (1990), Bakes and Nichols (1995), Kayama et al. (1971) является селахиловый спирт (18:1n-9) и его содержание колеблется от 40% до 76%. При сравнении состава алкильной цепи в АДАГ жира печени и тела у катрана Scualus acanthias (Malins et al., 1965) показало, что жир тела имеет более высокие количества 16:0 по сравнению с жиром печени. Наибольшее отличие хрящевых видов рыб и от моллюсков по 16:0 как главного спирта в жире тела наблюдается у кальмара Berryteuthis magister и зоопланктона Clione limacine (Hallgren et al., 1974a). У млекопитающих в составе АДАГ преобладают 18:0 и 18:1, которые составляют до 80% от общего количества спиртов (Таблица 1).
Исследования показали, что состав алкильной части в АДАГ и в фосфолипидах с простой эфирной связью, как правило сходен, что может служить свидетельством единого пути биосинтеза алкил-содержащих липидов (Wykle et al., 1979). Тем не менее, в некоторых работах показано, что жирные acyl-CoA редуктазы, роль которых - синтез спиртов из соответствующих ЖК идет быстрее с 16:0, 18:0 и 18:1, чем с насыщенными короткоцепочечными жирными кислотами (ЖК) или с полиненасыщенными жирными кислотами (ПНЖК) (Reichwald-Hacker, 1983; Hartvigsen et al., 2006).
Алкильные липиды и плавучесть морских животных
Кроме присутствия в клеточных мембранах и хранения энергии, большое содержание АДАГ в печени некоторых глубоководных акул, как полагают, играет существенную роль в поддержании нейтральной плавучести при вертикальных перемещениях. Различные глубоководные виды акул, например семейства Squaliformes, накапливают большие количества имеющих малую плотность липидов типа сквалена (плотность 0.86 г/мл) и АДАГ (плотность 0.89 г/мл) в печени. Сквален и АДАГ обеспечивают на 80% и 14% большей подъемной силы в морской воде, чем триглицериды (плотность 0.92г/мл), который является наиболее распространенной формой хранения липидов у животных. Malins и Barone (1970) показали, что АДАГ активно обменивается жире печени акулы S. acanthias, и предположили, что метаболизм АДАГ был связан с ролью печени как гидростатического органа. Они искусственно увеличили вес тела S. acanthias акулы и через 50 часов сравнили содержание ТГ и АДАГ в печени и в других органах. Результаты показали, что количество АДАГ быстро и значительно увеличивалось во всех органах акулы. Поскольку у S. acanthias отсутствует плавательный пузырь, то синтез АДАГ является компенсаторным механизмом в контроле плавучести акулы.
Биологическая активность алкильных липидов
Исследования биологической активности алкил-содержащих липидов начались сравнительно недавно. Брохульт и Холмберг (Brohult et al., 1954; 1963) используя неомыляемую часть жиров костного мозга теленка в качестве препарата содержащего AГ, наблюдали ускорение созревания белых кровяных клеток в процессе курса радиотерапии при детской лейкемии. Эта работа показала, что АГ можно использовать при лучевой терапии различных злокачественных опухолей (Brohult et al., 1977). При этом у пациентов принимавших АГ снижение количества лейкоцитов и тромбоцитов в крови было менее выражено и частота появления постлучевых свищей снижалась более чем в два раза. Именно эти работы дали начало многочисленным медико-биологическим исследованиям АГ.
Спектр биологической активности нейтральных алкильных липидов разнообразен и, это связывают с высокой метаболической активностью АГ. Уже в ранних работах было показано необычное распределение алкильных липидов в клетках злокачественных опухолей. Анализ на содержание 1-O-алкил- и 1-алкил-1-енильных- эфиров в нейтральных и полярных липидах в 17 видах злокачественных опухолей человека показал, что в опухолевых клетках, как правило, содержание АГ было большим, чем в здоровых клетках из тех же органов (Snyder and Wood, 1969). Аналогичные результаты получены на животных, где обнаружили более высокие уровни простых эфиров, и особенно нейтральных, в привитых опухолях мышей и крыс, чем в здоровых тканях (Snyder and Wood, 1968). В клетках гепатоклеточной карциномы человека уровень АГ был в среднем выше, чем в ткани без опухоли или в здоровой печени, при этом было отмечено необычное распределение индивидуальных АГ. Так, химиловый спирт доминировал в карциноме и соотношения между самым распространенным АГ, химиловым, батиловым и селахиловым спиртами в опухолевых клетках составляло 2:1:1, в то время как в нормальных клетках это соотношение было 1:2:2 (Lin et al., 1978). Высокие уровни в содержания холин-плазмалогена отмечены в злокачественных тканях легких по сравнению со здоровой тканью (Merchant et al., 1991) и в различных опухолях мозга (Albert and Anderson, 1977). С другой стороны, у пациентов с острой миелогенной лейкемией уровень плазманил-ФХ в холиновой фракции ФЛ составлял 10-20% по сравнению с 42% в нормальных полиморфоядерных лейкоцитах (Chabot и et al., 1990). Кроме того, уровни плазманил-ФХ в лейкемических клетках положительно коррелировали со степенью клеточной дифференцировки. В связи с выше изложенными фактами в 80-х годах ХХ века популярной была идея использовать алкильные липиды в качестве маркеров злокачественных опухолей. Однако более поздние работы по анализу нейтральных и полярных простых эфиров опухолевых клеток не позволили сделать однозначных выводов о роли алкильных производных в раковых клетках. По этой причине не возможно использовать относительное увеличение уровня простых эфиров при диагностике рака. Однако очевидно, что в опухолевых клетках происходят изменения связанные с метаболизмом простых эфиров и, прежде всего, с незначительным увеличением уровней 1-O-алкил- и 1-алкил-1-енильных эфиров глицерина и состава ЖК в sn-2 положении алкильных фосфолипидов. Вероятно, это является следствием изменений в активности ферментов биосинтеза простых эфиров в опухолевых клетках, которые зависят от вида опухоли и являются защитной реакцией клетки на не контролируемый рост раковой опухоли (Magnusson and Haraldson, 2011).
Алкильные липиды и вторичные мессенджеры
Как показано на Рис. 2 биосинтез нейтральных АДАГ сопряжен с биосинтезом 1-О-алкил-глицеро-фосфолипидов на стадии образования 1-О-алкил-2-ацилглицерола в клетках, таких как в моноцитоподобные клетки линии THP-1 (Hichami et al., 1997), эндотелиальных клетках (Marigny et al., 2002) или тромбоцитах (Pédrono et al., 2004). В эндотелиальных клетках 1-О-алкил-глицеро-фосфолипиды под действием фосфолипазы С трансформировались в 1-О-алкил-2-ацил-глицерол (Marigny et al., 2002), аналога диглицерида — диацил-глицерола, являющегося ингибитором протеинкиназы С. В моноцитоподобных клетках линии THP-1, под действием фосфолипазы А2 образуется 1-О-алкил-глицеро-фосфатидилхолин, предшественник фактора активации тромбоцитов ФАТ (Hichami et al., 1997). Увеличение включения 1-О-алкил-2-ацил - фосфатидилхолина в мембранные фосфолипиды, также способствует увеличению производства активных аналогов ФАТ при окислении ненасыщенных ЖК в 2-позиции глицерина (Kamido et al., 2002; Hartvigsen et al., 2006).
Алкильные липиды и индуцируемая активность сперматозоидов.
Сперматозоиды продуцируют ФАТ, представителя полярных алкильных липидов, и этот посредник играет важную роль в физиологии спермы (Kraus et al., 1994; Fukuda et al., 1994) и, поскольку АДАГ является предшественником ФАТ, то опосредовано может оказывать действие на функции спермы. Действительно, в результате выдерживания спермы in vitro со смесью природных АГ возростала подвижность, выживаемость и скоростные параметры сперматозоидов. Кроме того, при такой обработке сперма обладает лучшей способностью к оплодотворению при искусственном осеменении (Cheminade et al., 2002), и это нашло применение для многих видов млекопитающих. In vivo исследования показали, что пероральный прием 40г/день жира акулы или 1гр. чистых АГ в течении 28 дней достоверно улучшает подвижность и выживаемость спермы свиней (Mitre et al., 2004).
Нативный жир акулы, в качестве пищевой добавки (32г/сутки) вводили в диету 12 беременным и кормящим свиноматкам для определения влияние на рост и иммунный статус потомства (Mitre et al., 2005). После приема жира свиноматки имели более высокий уровень эритроцитов, гемоглобина в крови и возрастали концентрации IgG, АГ и ω-3 ПНЖК в молоке. У поросят также повысились лейкоциты и уровень иммуноглобулина IgG. Жир акулы в качестве пищевой добавки (вид не указан) супоросным и лактирующим свиноматкам индуцировал положительное влияние на иммунный статус помета и это связывают повышением доли АДАГ в молоке свиноматок потомства (Mitre et al., 2005).
Алкильные липиды и иммунитет
Нейтральные и полярные алкил-содержащие липиды значительно активизируют цитотоксические макрофаги, что приводит к повышению Fc-рецепторного фагоцитоза естественных киллеров, макрофагов, нейтрофилов и повышают гуморальный иммунный ответ и отдаляют реакцию гиперчувствительности (Berdel et al., 1980). Показано, что батиловый и селахиловый спирты стимулируют гемапоэз, эритропоэз, тромбоцитоз и гранулоцитоз в костном мозге животных после радиационного облучения (Edlund, 1954; Osmond et al., 1963). Влияние диеты с различными концентрациями природных АГ в диапазоне от 10, 50 и 250 пикомоля было исследовано на лактирующих крысах в связи с развитием иммунных реакций у потомства. После 10 дней кормления молоком у потомства значительно повысилось количество периферийных гранулоцитов и уровень иммуноглобулинов IgG и IgM в плазме. Это исследование показывает, что АГ в крысином молоке играют важную роль в развитии иммунного ответа у новорожденных крысят (Oh and Jandhav, 1994). Однако при обработке in vitro перитональных клеток самок мышей синтетическим додецилглицеролом в течении 30 минут было отмечено значительное увеличение количества IgG, в то время как уровень IgM не изменялся. При этом культивирование макрофагов с АГ не повышала их активности (Homma et al., 1990)
Yamamoto et al., (1988) отметили, что иньекция в кровь небольших количеств (10-100 нг) синтетического додецил-глицеролом (С-12) мышам in vivo приводило к возрастанию Fc-рецепторной опосредованной активации макрофагов уже на 5-й день приема, причем наиболее эффективной дозой препарата был прием 5 нг/кг живого веса животных. Введение батилового спирта в еще более низких концентрациях вызывало аналогичную активацию макрофагов. Это исследование подчеркивает, что положительное влияние АГ, как химиотерапевтических агентов, стимулирует через Fc- рецептор опосредованную фагоцитарную активность макрофагов.
Исследовали роль суммарных алкил-глицеридов выделенных из акульего жира (состав 14:0 – 0,7%; 16:0 – 9,1%; 16:1n-7 – 12,5%; 18:1n7 – 4,8%; 18:1n-9 – 68,1%) на кальций сигнальную систему в Jurkat клетках (культура клеток линии Т-лимфоцитов клеток для исследования острого лейкоза, T клеточной сигнальной системы, а также для обнаружения различных рецепторов хемокинов восприимчивых к вирусным заболеваниям, в частности ВИЧ). В эксперименте, в без кальциевом буфере, чистые АГ повышали внутриклеточную концентрацию Са+2, доказывая, что АГ способно индуцировать переток Са+2 через открытие кальциевых каналов. Это исследование показало, что верапамил, который является ингибитором кальциевых каналов L-типа не оказывает эффекта на транспорт кальция при введении АГ. АГ частично снимает ингибирование N-конотоксина, который является блокатором N-типа и активирует перенос Ca2+ (до 39% максимальной скорости), через измение мембранного потенциала. Таким образом, представлены доказательства, что АГ увеличивает приток Ca2+ в Jurkat клетках, возможно через модулирование проницаемости кальциевых каналов (Pedrono et al., 2004).
Tchorzewski et al., (2002) показали, что поддерживающее лечение компонентами жира печени акулы нормализует уровень клеток природных киллеров, продуцируя активные формы кислорода необходимого для образования лейкоцитов в периферической крови у людей страдающих активной формой ревматоидного артрита.
Pamblad et al., (1990) оценивали в какой степени различные чистые АГ могут инициировать функциональный ответ нейтрофилов. В эксперименте были использованы различные, как нейтральные, так и полярные алкильные липиды. ФАТ был самым мощным по способности образовывать оксидативный ответ, затем следовали синтетический 1-0-гексадецил-2-метокси-глицеро-3-ФХ; лизо-ФАТ, батиловый- и химиловый спирты проявляли только слабую активность; ФАТ был также наиболее активным липидом существенно повышающим уровень внутриклеточного Са+2. Авторы заключили, что все используемые в эксперименте алкильные липиды стимулировали функциональный ответ нейтрофилов ФАТ показал в равных условиях наибольший потенциал из всех исследованных этерных липидов как в кальциевом ответе, так и функциональном ответе нейтрофилов. Некоторые другие этерные холин-содержащие ФЛ повышали активность кальциевых каналов, но не давали оксидативного эффекта.
Интересное исследование по влиянию препарата «Bioмarin 570» из жира акулы (50% нейтральных АДАГ) на уровень C1q, CD4/CD8, факторов показывающих иммунный статус организма, было проведено Tchorzewski et al., (2005). Пациенты с ревматоидным артиритом получали по 3 г/сутки Bioмarin 570 в течении 30 дней. «Bioмarine 570» повышал уровень C1q, CD4/CD8 от 1,3 до 1,8 раза. При этом увеличивалась поляризация лимфоцитов, секреция цитокинов Th1 и наблюдалось активное образование нейтрофилов. Исследование не показало, никаких побочных эффектов, доказав что «Bioмarin 570» является безопасным, эффективно поддерживающим врожденный иммунитет препаратом и может быть использован для людей с нарушенной иммунной системой как безвредный и эффективный агент для приведения в норму иммунного дисбаланса.
Алкильные липиды в онкологии
Одно из наиболее интересных свойств АГ оказалась их противоопухолевая активность. Сравнительное исследование приема чистых природных нейтральные АГ и жира печени акулы per os мышами с карциномой легких Льюиса показало, что чистые АГ снижали метастазирование на 64 ± 8%, тогда как эффект от жира составил 30 ± 9% по сравнению с контролем. Очищенные AГ также снизили общее содержание плазмалагенов в опухолях, в то время как с жир не показал какого либо эффекта. На 5-й день приема AГ понизился в опухоли фактор фон Виллебранда, маркер эндотелиальных клеток, что свидетельствует об антиангиогенном действии AГ. Это исследование показало, что чистые AГ снижают скорость васкуляризации и распространение рака опухолей у мышей (Pedrono et al., 2004).
В другой работе выполненной Hajimoradi et al. (2009) по противораковому эффекту очищенного акульего жира с 10% содержанием АДАГ и витаминами A, D, Е животным вводили внутрибрюшинно различные количества жира - 50, 10, 5, 2,5 и 0,1 мкг/кг/сут. Эксперименты показали, что, дозы 50 и 10 мкг/кг/сут, дали максимальный эффект на гиперчувствительность замедленного типа уже после 48 часов, увеличивалось количество CD8+ лимфоцитов и образование гамма-интерферона, при этом наблюдалось снижение скорости роста опухоли. При введении 5 и 2,5 мкг/кг/сут снижение скорости роста опухоли проявлялось слабо. Эти результаты подчеркивают, что акулий жир может быть использован для профилактики и лечения раковых больных с ослабленным иммунитетом.
Krotkiewski et al. (2003) для оценки влияния AГ использовал клеточные линии карциномы яичников человека (OVP-10), рака молочной железы (MCF-7) и 3 клеточных линии рака простаты (DU-145, PC-3 и PCA-2b). Клетки обрабатывались препаратом «Ecomer», содержащим 20% AГ и 3% метокси производных нейтральных АГ в дозе от 0,1 мг/мл до LD-50. Клетки простаты DU-145, PC-3 и PCA-2B показали значительное увеличение апоптозных клеток даже при небольших дозах - 0,5 и 0,1 мг/мл, в то время как в клетках рака молочной железы отмершие клетки появлялись после достаточно высоких концентраций препарата. Это исследование показывает, что процессы апоптоза/некроза вызванные «Ecomer» у трех дифференцированных клеточных линий рака простаты, молочной железы и клеток карциномы проходят с высокой скоростью. Однако, такого эффекта не наблюдалось на клетках рака яичников.
В сравнительном исследовании эффекта чистых АГ (из жира печени акулы) и нативного жира печени акулы (содержащий 25% АДАГ без сквалена) на рост опухоли, легочные метастазы и васкуляризации клеток карциномы легкого Льюиса (3LL) привитой мышам было показано, что и жир и чистые АГ уменьшали 3LL-привитую карциному на 29% и 26%, соответственно, по сравнению с контролем. Лечение жиром печени акулы вызвало 31%-ое снижение легочных метастаз, в то время как нейтральные АГ снижали количество метастаз на 64%. Интересно, что обработка АГ вызвала существенное сокращение кровеносных сосудов в опухоли, поэтому противоопухолевый эффект АГ происходит, по крайней мере частично, Авторы обьясняют их антиангиогенной активностью в клетках опухоли (Pedrono et al., 2004). Однако противораковая активность зависела от состава спиртовой группы. Так селахиловый (16:0) и 16:1n-7 АГ проявили самую высокую степень подавления роста опухоли и cнижение числа метастаз у мышей, привитых 3LL клетками, по сравнению с 16:0 и 12:0 АГ (Deniau et al., 2010).
Pedrono et al., (2007) исследовали влияние чистых нейтральных AГ (14:0 - 0,7%, 16:0 - 9,1%, 16:1n-7 = 12,5%, 18:1 н-9 = 68,1%, 18:1 н -7 = 4,8% и другие (<1%) = 4.8%) на пролиферацию основного фактора роста фибробластов (bFGF) стимулированных эндотелиальных клеток. Нейтральные AГ метаболизировали в несколько липидов, которые могли влиять на передачу сигнала, а именно в алкил-фосфатидную кислоту и ее лизопроизвоное, алкил-фосфатидил-холин, алкил-фосфатидилинозит и алкил-моноацил-глицерин. AГ влиял на рост клеток эндотелия, без цитотоксических эффектов, снижал пролиферацию клеток в зависимости от концентрации и времени через ингибирование фактора bFGF.
Интересные результаты были полученыи несколькими группами исследователей при тестировании нейтральных метокси-производных АГ. Так Wang et al., (1999) показали, что метокси-АГ могут угнетать доброкачественные или дифференцированные фенотипы клеточных линий трех видов рака толстой кишки человека. Метокси-АГ ингибировали рост всех трех типов клеток в одинаковой степени, 80% ингибирования роста клеток наблюдалось при 25 мМ. При этом повышалось образование эмбрионального антигена во всех трех клеточных линиях и продуцировался раковый эмбриональный антиген (CEA) в к-ве 5 ng/106 клеток. Повышение образования CEA наблюдается и в клетках обработанных 10, 25 или 50 мМ метокси-АГ. Максимальная индукция CEA (8 ng/106 клеток) наблюдалось, когда клетки обрабатывали 25 мкМ метокси-АГ. Кроме того, в работе показано, что метокси-АГ снижают порог чувствительности раковых новообразований при радиотерапии и могут использоваться в качестве сопутствующего эффективного метода при лечении рака.
Аналогичные результаты были получены при обработке 2 клеточных линий рака простаты человека LNCaP и DU145 М-АГ (Reynolds et al., 2000). В этом исследовании метокси-АГ ингибировали рост клеток в равной степени с IC-50 от 93 мкМ для LNCaP и 97 мкМ для DU145, в то время как IC-50 значения для фенилбутирата были 1,3 мМ и 7,3 мМ соответственно, для тех же клеток. Метокси-АГ угнетают пролиферацию, прикрепление и рост обеих линий клеток простаты человека. Авторы связывают противоопухолевый эффект метокси-АГ с активацией макрофагов, а также с увеличением продукции цитокинов, таких как интерлейкина-12 (IL-12) и гамма-интерферона. Считается, что увеличение количества и активация макрофагов являются основой в первичной противоопухолевой защите, в то время как γ-интерферон и Т-клетки продуцирующие лимфокины, тормозят многие виды раковых клеток, либо непосредственно или препятствуют росту раковых клеток через иммуномодулирующие свойства (Ucer et al., 1987). Другой возможный сценарий, объясняющий противоопухолевую активностью AГ может заключаться в накоплении О-алкильных групп в опухолевых клетках имеющих низкую О-алкил монооксигеназную активность, что приводит к накоплению этерных липидов в мембранах клеток и, в конечном итоге, к гибели клеток (Hoffman et al., 1984).
АГ гематоэнцефалический барьер
Гематоэнцефалический барьер (ГЭБ) выполняет функцию высокоселективного фильтра, через который из кровеносного русла в мозг поступают питательные вещества, а в обратном направлении выводятся продукты жизнедеятельности нервной ткани. Вместе с тем, наличие ГЭБ затрудняет лечение многих заболеваний центральной нервной системы, так как представляет серьезное препятствие на пути доставки лекарственных препаратов в центральную нервную систему.
Дифференциальная проницаемость ГЭБ в отсутствие и в присутствии 200 мМ 1 – О-пентил-глицерола исследованы как на безопухолевых крысах, так с животными с привитой С6 глиомой. В качестве лекарственного препарата был использован метатрексат, препарат использующийся при химиотерапии рака. В качестве маркеров AГ-опосредованного открытия ГЭБ были использованы флуоресцентные метки. Исследования показали, что 1-О-пентилглицерол увеличивает вход метатрексата в ткани опухоли головного мозга. (Erdlenbruch et al., 2003а). В другой работе (Erdlenbruch et al., 2003b) на здоровых самцах крыс была показана эффективность и зависимость преодоления ГЭБ от структуры нейтральных АГ. Наиболее перспективными являются пентил- и гексил-глицеролы. Эффекты были сравнены с нарушениями ГЭБ при использовании гипертонического маннита или введением в сонную артерию брадикинина. Авторы отмечают, что все AГ выделенные из природных обьектов вызывали увеличение проницаемости для метатрексата в мозг в зависимости от концентрации до 300 раз выше по сравнению с внутриартериальным введением только метотрексата. Однако повышение ГЭБ быстро подходит к исходному уровню (от 5 до 120 мин), что связывают со скоростью метаболизма АГ. В экспериментах на культурах клеток и при введении в сонную артерию животных не было выявлено каких-либо признаков токсичности. Таким образом, введение в сонную артерию АГ представляет новые терапевтические возможности для преодоления ограниченного доступа лекарств в центральную нервную систему.
Антимикробная и антибактериальная активность алкил-глицеридов.
Простые эфиры жирных кислот с многоатомными спиртами известны как широко распространенные антимикробные агенты, среди которых монолаурин был известен как наиболее активный (Kabara, 1979; Kabara et al., 1977). Однако недавно было показано, что додецил-АГ имеет значительно более высокий потенциал из-за высокой метаболической и химической стабильности этерной связи. Считается, что додецил-АГ оказывает антибактериальное действие через высвобождение и активацию протеаз и ингибирование синтеза пептидогликана в клеточных стенках бактерий. Brissette et al., (1986) использовали Streptococcus mutans BHT для изучения ответа эфиров додецил-АГ на ингибирование роста. S. mutans BHT устойчив к пенициллину и ряду других клеточных ингибиторов на основе антибиотиков, но все они стимулируют синтез липидов, тем самым блокируют клеточный лизис. Додецил-АГ ингибирует включение глицерина и биосинтез липидов S. mutans BHT при концентрациях 10 - 20мкг/мл, блокируя развитие S. mutans.
Haynes et al., (1994) тестировали бактерицидные и противогрибковые свойства додецил-АГ по отношению к дрожжам Candida и Cryptococcus, которые являются основными родами вызывающие грибковые инфекции у больных СПИД. Всего было исследовано действие АГ на Cryptococcus neoformans (6 штаммов), Cryptococcus albidus (1 штамм) и Cryptococcus laurentii (1 штамм). Из рода Candida, Candida аlbicans (1 штамм), Candida tropicalis (2 штамма,) и Candida parapsilosis (2 штамма). Авторы показали, что додецил-АГ ингибировал рост всех видов дрожжей Candida и Cryptococcus и эффект был синергетичен с амфотерицином В, при этом додецил-АГ снижал минимальную ингибирующую концентрацию амфотерицина в 80 раз. Другие АГ с длиной цепи от С-8 до С-18 также проявляли синергизм с амфотерицином. Однако нейтральные АГ не проявляли синергизма с с водо-растворимыми антигрибковым сединением флюконазолом, подтверждая, что гидрофобные свойства амфотерицина В играют ключевую роль в синергизме.
Влияние алкил-глицеридов на протеинкиназу C
Протеин киназа C (ПКС) играет важную роль как медиатор клеточных ответов на внеклеточные стимулы, то есть осуществляет трансдукцию сигнала. Эта функция, имеет большую значимость для различных клеточных процессов, включая регулирование клеточной адгезии, рост и дифференцировку клеток (Dekker and Parker, 1994; Nishizuka, 1986). Известно, что существует прямая корреляция между способностью эфиров форбола избирательно активировать ПKC и вызывать образование опухолей (Nishizuka, 1984). Отмечены высокие уровни ПКС в клетках опухоли молочных желез, легких и желудка и поэтому предполагется, что некоторые ПKC-изоформы могут быть потенциальными целями при лечении некоторых видов рака (Mackay и Twelves, 2003). ПКС - семейство серин/треониновых киназ, состоящих, по крайней мере, из 12 изоформ, функция которых заключаются в активации через фосфорилирование некоторых белков, вовлеченных в различные процессы клеток. Активация определенного рецептора внеклеточных сигнальных молекул связана с активацией фосфолипазы C которая, в свою очередь, гидролизует мембран-связанный фосфатидилинозит-4,5-бисфосфат, образуя два вторичных мессенджера - 1,2-диацил-глицерол (ДАГ), который остается в мембране клеток, и инозитол-1,4,5-трифосфат (ИФ3), который уходит в цитозоль. ИФ3 стимулирует освобождение ионов Ca2+ из эндоплазматического ретикулума в цитозоль и вызывает перемещение неактивной цитозольной ПКС в цитоплазму, где она активируется ДАГ (Heymans et al., 1987; Alberts, 1994). ДАГ играет центральную роль в ПКС регулировании роста клеток, и показывает, что его концентрация повышается в пролиферативных клетках (Warne et al., 1995). Нейтральные АГ являются ингибиторами ПКС, поскольку в 1 положении глицерина вместо ацильной группы находится алкильный радикал. В культуре клеток почки собаки, нейтральные АГ вызвали ингибирование ПКС. 1-O-dodecyl-АГ снижал активность ПКС в не стимулируемых клетках и блокировал активацию ПКС эфирами форбола. Кроме того, АГ активно влияли на скорость роста клеток и накапливались в клетках почек собаки, пролонгируя эффект ингибирования (Warne et al., 1995).
Ингибирующие свойства чистых АГ были показаны на очищенной ПКС с чистыми синтетическими 1-гексадецил- и 1-O-октадецил АГ и со смесью природных АГ, имеющих 1-O-алкильные цепи до С-22 и С-24 (Warne et al., 1995). Интересно, что ацильный аналог химилового спирта, то есть 1-монопальмитоил-глицерин, проявил очень низкую ингибирующую способность к ПКС а диалкил-глицерид 1,2-O-гексадецил-глицерин не влиял на активность ПКС (McNeely et al., 1989).
Ингибирование ПКС нейтральными АГ и алкенильными аналогами фосфолипидов, было показано на эндотелиальных клетках аорты свиньи (Marigny et al., 2002). На основе проведенных экспериментов был сделан вывод, что антираковые св-ва АГ могли бы быть следствием ингибирования некоторых ПКС изоформ в клетках опухоли, поскольку различные ПКС изозимы вовлечены в метастазирование опухоли. Действительно, было показано, что ингибирование ПКС-изоформ препятствовало развитию опухоли в эндотелиальных клетках у мышей (Yoshiji et al., 1999). Ранее отмечали, что именно нейтральные АГ выделенные из жира печени акул были ответственны за анти-ангиогенные свойства жира печени акулы в клетках карциномы легкого Льюиса у мышей (Pedrono et al., 2004). Кроме того, то же самое исследование показало, что АГ увеличил проводимость эндотелиальных клеток, возможно приводя к изменениям в цитоскелете и при слипании клеток, что могло приводить к ингибированию роста опухоли.
Литература
Куликов В.И., Музя Г.И. Алкоксиглицеролипиды и фактор активации тромбоцитов: эволюция и функция в клетке. Биохимия (Россия), 1997, 62(10), 1288-1294.
Alberts, B., Bray, D., Lewis, J., Raff, M., Roberts, K., Watson, J.D. 1994. Cell signaling. In: Alberts, B., Bray, D., Lewis, J., Raff, M., Roberts, K., Watson, J.D. (Eds.), Molecular Biology of the Cell., 3rd ed. Garland Publishing, New York, pp. 721–785.
Albert, D.H., Anderson, C.E. Ether-linked glycerolipids in human-brain tumors. Lipids 1977, 12, 188–192.
Bakes, M.J., Nichols, P.D. Lipid, fatty acid and squalene composition of liver oil from six species of deep-sea sharks collected in southern Australian waters. Comp. Biochem. Physiol. B 1995, 110, 267–275.
Benveniste, J., Vargaftig, B.B. 1983. Platelet-activating factor: an ether lipid with biological activity. In: Mangold, H.K., Paltauf, F. (Eds.), Ether Lipids: Biochemical and Biomedical Aspects. Academic Press, New York, pp. 355–376.
Berdel, W.E.; Bausert, W.R.; Weltzien, H.U.; Modotell, M.L.; Widmann, K.H.; Munder, P.G. The influence of alkyl-lysophospholipids and lysophospholipid activated macrophages on the development of metastasis of 3-Lewis lung carcinoma. Eur. J. Cancer 1980, 16, 1199–1204.
Brissette, J.L.; Erlinda, A.; Cabacungan, E.A.; Pieringer, R.A. Studies on the Antibacterial Activity of Dodecylglycerol. Its limited metabolism and inhibition of glycerolipid and lipoteichoic acid biosynthesis in Streptococcus Mutans BHT. J. Biol. Chem. 1986, 261, 6338–6345.
Brites, P., Waterham, H.R., Wanders, R.J.A. Functions and biosynthesis of plasmalogens in health and disease. Biochim. Biophys. Acta 2004, 1636, 219–231.
Brohult, A., Holmberg, J. Alkylglycerols in the treanment of leucopenia caused by irradiation. Nature 1954, 174, 1102-1103.
Brohult, A. Alkylglycerols and their use in radiation treatment. Acta Radiol. Ther. Phys. Biol. 1963, 24, ( Suppl. 223), 1-99.
Brohult, A., Brohult, J., Brohult, S., Joelsson, I. Effect of alkylglycerols on the frequency of injuries following radiation therapy for carcinoma of the uterine cervix. Acta Obstet. Gynecol. Scand. 1977, 56, 441-448.
Brohult, A., Brohult, J., Brohult, S., Joelsson, I. Effect of alkylglycerols on the frequency of fistulas following radiation therapy for carcinoma of the uterine cervix. Acta Obstet. Gynecol. Scand. 1979, 58, 203-207.
Chabot, M.C., Greene, D.G., Brockschmidt, J.K., Capizzi, R.L., Wykle, R.L. Etherlinked phosphoglyceride content of human leukemia cells. Cancer Res. 1990, 50, 7174–7178.
Cheminade, C., Gautier, V., Hichami, A., Allaume, P., Le Lannou, D., Legrand, A.B. 1-O-Alkylglycerols improve boar sperm motility and fertility. Biol. Reprod. 2002, 66, 421–428.
de Vet, E.C.J.M., Hilkes, Y.H.A., Fraaije, M.W., van den Bosch, H. Alkyldihydroxyacetone phosphate synthase. Presence and role of flavin adenine dinucleotide. J. Biol. Chem. 2000, 275, 6276–6283.
Dekker, L.V., Parker, P.J. Protein kinase C—a question of specificity. Trends Biochem. Sci. 1994, 19, 73–77.
Deniau, A.L., Mosset, P., Pedrono, F., Mitre, R., Le Bot, D., Legrand, A.B. Multiple beneficial health effects of natural alkylglycerols from shark liver oil. Mar. Drugs 2010, 8, 2175–2184.
Deprez, P.P., Volkman, J.K., Davenport, S.R. Squalene content and neutral lipid composition of livers from deep-sea sharks caught in Tasmanian Waters. Aust. J. Mar. Fresh. Res. 1990, 41, 375–387.
Edlund, T. Protective effect of d,1-alfa-octadecylglycerol ether in mice given total body x-irradiation. Nature 1954, 174, 1102.
Erdlenbruch, B., Alipour, M., Fricker, G., Miller, D.S., Kugler, W., Eibl, H., Lakomek, M. Alkylglycerol opening of the blood-brain barrier to small and large fluorescence markers in normal and C6 glioma-bearing rats and isolated rat brain capillaries. Brit. J. Pharmacol. 2003a, 140, 1201–1210.
Erdlenbruch, B., Jendrossek, V., Eibl, H., Lakomek, M. Transient and controllable opening of the blood-brain barrier to cytostatic and antibiotic agents by alkylglycerols in rats. Exp. Brain Res. 2003b, 135, 417–422.
Fukuda, A.; Roudebush, W.E.; Thatcher, S.S. Platelet-activating factor enhances the acrosome reaction, fertilization in vitro by subzonal sperm injection and resulting embryonic development in the rabbit. Hum. Reprod. 1994, 9, 94–99.
Hajimoradi, M.; Daneshmandi, S.; Hassan, Z.M. Effect of Shark Liver Oil on Nitric Oxide Production and MTT Reduction by Peritoneal Macrophages from BALB/c mice. Available online: http://www.mums.ac.ir/shares/research/sazagarf1/64-70(41).pdf (Accessed on 19 June
Hallgren, B., Niklasson, A., Stallberg, G., Thorin, H. On the occurrence of 1-O-(2-methoxyalkyl)glycerols and l-O-phytanylglycerol in marine animals. Acta Chem. Scand. B 1974a, 28, 1035–1040.
Hallgren, B., Niklasson, A., Stallberg, G., Thorin, H. On the occurrence of 1-Oalkylglycerols and 1-O-(2-methoxyalkyl)glycerols in human colostrum, human milk, cow’s milk, sheep’s milk, human red bone marrow, red cells, blood plasma and a uterine carcinoma. Acta Chem. Scand. B 1974b, 28, 1029–1034.
Hartvigsen, K., Ravandi, A., Harkewicz, R., Kamido, H., Bukhave, K., Holmer, G., Kuksis, A. 1-O-Alkyl-2-(-oxo)acyl-sn-glycerols from shark oil and human milk fat are potential precursors of PAF mimics and GHB. Lipids 2006, 41, 679–693.
Heymans, F.; Da Silva, C.; Marrec, N.; Godfroid, J.-J.; Castagna, M. Alkyl analogs of diacylglycerols as activators of protein kinase C. FEBS Lett. 1987, 218, 35–40.
Haynes, M.P.; Buckley, H.R.; Higgins, M.L.; Pieringer, R.A. Synergism between the Antifungal Agents Amphotericin B and Alkyl Glycerol Ethers. Antimicrob. Agents Chemother. 1994, 38,1523–1529.
Hichami, A., Duroudier, V., Leblais, V., Vernhet, L., Le Goffic, F., Ninio, E., Legrand, A. Modulation of platelet-activating-factor production by incorporation of naturally occurring 1-O-lkylglycerols in phospholipids of human leukemic monocyte-like THP-1 cells. Eur. J. Biochem. 1997, 250, 242–248.
Hoffman, D.R.; Hadju, J.; Snyder, F. Cytotoxicity of PAF and related alkyl-phospholipid analogs in human leukaemia cells, polymorphonuclear neutrophils, and skin fibroblasts. Blood 1984, 63, 545–552.
Homma, S.; Millman, I.; Yamamoto, N. A serum factor for macrophage activation after in vitro dodecylglycerol treatment of mouse lymphocytes. Immunol. Cell Biol. 1990, 67, 137–142.
Iannitti, T., Palmieri, B. An update on the therapeutic role of alkylglycerols. Mar. Drugs 2011, 8, 2267–2300.
Jayasinghe, C., Gotoh, N., Wada, S. Variation in lipid classes and fatty acid composition of salmon shark (Lamna ditropis) liver with season and gender. Comp. Biochem. Physiol. B 2003, 134, 287–295.
Kabara, J.J. Fatty Acids and Derivatives as Antimicrobial Agents-A Review. In The Pharmacological Effect of Lipids; Kabara, J.J., Ed.; The American Oil Chemists' Society: Champaign, IL, USA, 1979, 1–14.
Kabara, J.J.; Vrable, R.; Lie Ken Jie, M.S.F. Antimicrobial lipids: Natural and synthetic fatty acids
and monoglycerides. Lipids 1977, 12, 753–759.
Kamido, H.; Eguchi, H.; Ikeda, H.; Imaizumi, T.; Yamana, K.; Hartvigsen, K.; Ravandi, A.; Kuksis, A. Core aldehydes of alkyl glycerophosphocholines in atheroma induce platelet aggregation and inhibit endothelium-dependent arterial relaxation. J. Lipid Res. 2002, 43, 158–166.
Kayama, M., Tsuchiya, Y., Nevenzel, J.C. Glyceryl ethers of some shark liver oils. B. Jpn. Soc. Sci. Fish. 1971, 37, 111–118.
Koga, Y., Morii, H. Recent advances in structural research on ether lipids from archaea including comparative and physiological aspects. Biosci. Biotech. Biochem. 2005, 69, 2019–2034.
Kossel, A., Edlbacher, S. Beitrage zur chemischen Kenntnis der Echinodermen. Z. Physiol. Chem. 1915, 94, 264–283.
Kraus, C.S.; Gervasi, G.; Fori, G.; Baldi E. Effect of platelet-activating factor on motility and acrosome reaction on human spermatozoa. Hum. Reprod. 1994, 9, 471–476.
Krotkiewski, M., Przybyszewska, M., Janik, P. Cytostatic and cytotoxic effects of alkylglycerols (Ecomer). Med. Sci. Monit. 2003, 9, 131–135.
Lin, H.J., Ho, F.C., Lee, C.L. Abnormal distribution of O-alkyl groups in the neutral glycerolipids from human hepatocellular carcinomas. Cancer Res. 1978, 38, 946–949.
Mackay, H.J., Twelves, C.J. Protein kinase C: a target for anticancer drugs? Endocr-Relat. Cancer 2003, 10, 389–396.
Magnusson, C.D., Haraldsson, G.G., Ether lipids. Chem. Phys. Lipids 2011, 164, 315-340.
Malins, D.C., Wekell, J.C., Houle, C.R. Composition of the diacyl glyceryl ethers and triglycerides of the flesh and liver of the dogfish (Squalus acanthias). J. Lipid Res. 1965, 6, 100–105.
Malins, D.C., Barone, A. Glyceryl ether metabolism. Regulation of buoyancy in dogfish Squalus acanthias. Science 1970, 167, 79–80.
Mangold, H., Malins, D.C. Fractionation of fats, oils, and waxes on thin layers of silicic acid. J. Am. Oil Chem. Soc. 1960, 37, 383–385.
Marigny, K., Pedrono, F., Martin-Chouly, C.A.E., Youmine, H., Saiag, B., Legrand, A.B. Modulation of endothelial permeability by 1-O-alkylglycerols. Acta Physiol. Scand. 2002, 176, 263–268.
McNeely, T.B., Rosen, G., Londner, M.V., Turco, S.J. Inhibitory effects on protein kinase C activity by lipophosphoglycan fragments and glycosylphosphatidylinositol antigens of the protozoan parasite Leishmania. Biochem. J. 1989, 259, 601–604.
Merchant, T.E., Meneses, P., Gierke, L.W., Den Otter, W., Glonek, T. 31P magnetic resonance phospholipid profiles of neoplastic human breast tissues. Brit. J. Cancer 1991, 63, 693–698.
Mitre, R.; Cheminade, C.; Allaume, P.; Legrand, P.; Legrand, A.B. Oral intake of shark liver oil modifies lipid composition and improves motility and velocity of boar sperm. Theriogenology 2004, 62, 1557–1566.
Nishizuka, Y. The role of Protein kinase C in cell-surface signal transduction and tumor promotion. Nature 1984, 308, 693–698.
Nishizuka, Y. Studies and perspectives of Protein kinase C. Science 1986, 233, 305–312.
Oh, S.Y.; Jadhav, L.S. Effects of dietary alkylglycerols in lactating rats on immune responses in pups. Pediatr. Res. 1994, 36, 300–305.
Osmond, D.G.; Roylance, P.J.; Webb, A.J.; Yoffley, J.M. The action of batyl alcohol and selachyl alcohol on the bone marrow of the guinea pig. Acta Haematol. 1963, 29, 180–186.
Paltauf, F. Metabolism of the enantiomeric 1-O-alkyl glycerol ethers in the rat intestinal mucosa in vivo; incorporation into 1-O-alkyl and 1-O-alk-I’-enyl glycerol lipids. Biochim. Biophys. Acta 1971, 239, 38–46.
Paltauf, F. 1983 Biosynthesis of 1-O-(1-alkenyl) glycerolipids (plasmalogens). In: Mangold, H.K., Paltauf, F. (Eds.), Ether Lipids: Biochemical and Biomedical Aspects. Academic Press, New York, pp. 107–128.
Pedrono, F.; Cheminade, C.; Legrand, A.B. Natural 1-O-alkylglycerols reduce platelet-activating factor-induced release of [3H]-serotonin in rabbit platelets. Prostaglandins Leukot. Essent. Fatty Acids 2004, 71, 19–23.
Pedrono, F., Martin, B., Leduc, C., Le Lan, J., Saiag, B., Legrand, P., Moulinoux, J.P., Legrand, A.B. Natural alkylglycerols restrain growth and metastasis of grafted tumors in mice. Nutr. Cancer 2004, 48, 64–69.
Pethybridge, H., Daley R., Virtue P., Nichols P. Lipid composition and partitioning of deepwater
chondrichthyans: inferences of feeding ecology and distribution. Mar. Biol. 2010, 157, 1367–1384
Phleger C.F., Nichols P.D., Virtue P. Lipids and buoyancy in soutern ocean pteropods. Lipids. 1997, 32, 1093-1100.
Reichwald-Hacker, I. 1983. Substrate specificity of enzymes catalyzing the biosynthesis of ether lipids. In: Mangold, H.K., Paltauf, F. (Eds.), Ether Lipids: Biochemical and Biomedical Aspects. Academic Press, New York, pp. 129–140.
Reynolds, S., Cederberg, H., Chakrabarty, S. Inhibitory effect of 1-O (2 methoxy) hexadecyl glycerol and phenylbutyrate on the malignant properties of human prostate cancer cells. Clin. Exp. Metastas. 2000, 18, 309–312.
Roudebush, W.E., Massey, J.B., Elsner, C.W., Shapiro, D.B., Mitchell-Leef, D., Kort, H.I., The significance of platelet-activating factor and fertility in the male primate: a review. J. Med. Primatol. 2005, 34, 20–24.
Snyder, F. The ether lipid trail: a historical perspective. Biochim. Biophys. Acta 1999, 1436, 265–278.
Snyder, F., Wood, R., The occurrence and metabolism of alkyl and alk-1-enyl ethers of glycerol in transplantable rat and mouse tumors. Cancer Res. 1968, 28, 972–978.
Snyder, F., Wood, R. Alkyl and alk-1-enyl ethers of glycerol in lipids from normal and neoplastic human tissues. Cancer Res. 1969, 29, 251–257.
Sprott G.D. Structure of archaebacterial membrane-lipids. J.Bioenerget.Biomembran. 1992, v.24, 555-566. DOI: 10.1007/BF00762348
Tchórzewski, H., Banasik, M.; Głowacka, E.; Lewkowicz, P. Modification of innate immunity in humans by active components of shark liver oil. Pol. Merkur. Lekarski 2002, 13, 329–332.
Tsujimoto, M., Toyama, Y. Ueber die unverseifbaren Bestandteile (hoheren Alkohole) der Haifishund Rochen-leberole. I. Chemische Umschau auf dem Gebiet der Fette, Oele, Wachse und Harze 1922, 29, 43-45
Wang, H., Rajagopal, S., Reynolds, S., Cederberg, H., Chakrabarty, S. Differentiation promoting effect of 1-O (2 methoxy) hexadecyl glycerol in human colon cancer cells. J. Cell. Physiol. 1999, 178, 173–178.
Warne, T.R., Buchanan, F.G., Robinson, M. Growth-dependent accumulation of monoalkylglycerol in Madin-Darby canine kidney cells. Evidence for a role in the regulation of Protein kinase C. J. Biol. Chem. 1995, 270, 11147–11154.
Wetherbee, B.M., Nichols, P.D. Lipid composition of the liver oil of deep-sea sharks from the Chatham Rise, New Zealand. Comp. Biochem. Physiol. 2000, B 125, 511–521.
Wykle, R.L., Malone, B., Snyder, F. Acyl-Coa reductase specificity and synthesis of wax esters in mouse preputial gland tumors. J. Lipid Res. 1979, 20, 890–896.
Yoshiji, H., Kuriyama, S., Ways, D.K., Yoshii, J., Miyamoto, Y., Kawata, M., Ikenaka, Y., Tsujinoue, H., Nakatani, T., Shibuya, M., Fukui, H. Protein kinase C lies on the signaling pathway for vascular endothelial growth factor-mediated tumor development and angiogenesis. Cancer Res. 1999, 59, 4413–4418.